DENTS
Situées à l’entrée du tube digestif, les dents constituent des organes très particuliers et hautement différenciés, tant par leur structure histologique très spéciale que par leurs remarquables propriétés mécaniques et physiologiques. En effet, d’un point de vue fonctionnel, on peut définir les dents comme des organes extrêmement durs, destinés à saisir, à retenir et à broyer les aliments.
Toutefois, l’étude détaillée des dents (et des tissus dentaires) révèle qu’elles ont une signification morphologique beaucoup plus générale: une définition purement fonctionnelle des dents inclurait sous ce terme des formations qui en remplissent les fonctions sans en avoir la structure et, réciproquement, une définition uniquement histologique les confondrait avec divers constituants de l’exosquelette (écailles, par exemple) des Vertébrés inférieurs. Pour parvenir à une définition satisfaisante, il est donc indispensable de faire appel à la fois aux critères histologiques et fonctionnels. Afin de comprendre la signification évolutive de ces formations, en apparence si isolées parmi les autres structures organiques, il est nécessaire également de faire appel aux ressources de l’anatomie et de l’histologie comparées, ainsi qu’à la paléontologie.
La conjonction de ces points de vue met en évidence les rapports fondamentaux et constants des tissus dentaires avec la peau et le squelette, rapports peu visibles dans une perspective limitée aux seuls points de vue classiques, notamment l’adaptation des dents au régime alimentaire chez les Mammifères.
1. Les tissus dentaires
Une dent de Mammifère, comme une écaille placoïde (fig. 1) de Sélacien, se forme à partir d’une ébauche mixte, épidermo-dermique (fig. 2). L’ébauche épidermique (d’origine ectodermique) différencie à la périphérie du germe de la dent des cellules spécialisées (adamantoblastes) qui assurent la sécrétion de l’émail, dont les couches se superposent du dedans au dehors (croissance centrifuge).
Recouverte par la précédente, l’ébauche dermique (d’origine mésodermique) différencie des cellules (odontoblastes) qui produisent la dentine (ou ivoire), tissu qui se dépose de dehors en dedans (croissance centripète).
Au centre de la dent, une cavité pulpaire entourée de dentine contient des vaisseaux sanguins et lymphatiques, ainsi qu’une ramification sensitive issue des subdivisions maxillaires ou mandibulaires du trijumeau. Enfin, au voisinage immédiat de la région radiculaire de la dent, le derme fibreux qui l’inclut se modifie et subit une calcification, donnant un tissu paradentaire, le cément.
L’émail que produisent les adamantoblastes, très fortement minéralisé (de 96 à 98 p. 100), forme des couches peu épaisses toujours avasculaires. Les adamantoblastes sont refoulés par la substance extracellulaire qu’ils produisent; celle-ci est donc anhiste mais peut contenir de fins canalicules. La substance organique extracellulaire est extrêmement réduite et la substance minérale est l’hydroxyapatite. Les cristaux de cette substance sont organisés typiquement en «prismes» de 4 à 6 猪m de large et striés longitudinalement (fig. 3).
L’émail des dents de Mammifères peut donc se définir par sa position superficielle (origine épidermique) et sa structure prismatique, mais aucun de ces critères n’est absolument applicable dans tous les cas chez les autres Vertébrés. On peut appeler «tissus adamantins» les tissus durs de certains Vertébrés inférieurs (ganoïne, vitrodentine, durodentine), qui se rapprochent de l’émail mammalien par l’aspect, la position et la composition, mais dont la signification n’est pas toujours clairement établie.
La dentine est un tissu dur minéralisé à 70 p. 100. La portion extracellulaire comprend une trame organique, principalement formée de fibrilles collagènes, et une fraction minérale formée d’hydroxyapatite. Les odontoblastes – et c’est une de leurs caractéristiques principales – ne sont jamais inclus entièrement dans la substance extracellulaire, contrairement aux ostéocytes de l’os. Seuls des prolongements cytoplasmiques sont présents dans des canalicules parallèles et branchus qui parcourent la portion extracellulaire du tissu. Le reste de la cellule se trouve continuellement repoussé par la substance extracellulaire qu’il produit: l’odontoblaste est donc une cellule polarisée (fig. 3). Cette description très générale s’applique à la dentine des Mammifères (orthodentine) et des Reptiles. Dans les autres classes de Vertébrés, sa structure peut différer notablement: la dentine peut être richement vascularisée (vasodentine) chez certains Téléostéens, repliée en lobes (plicidentine) chez les Labyrinthodontes et les Crossoptérygiens. Les ostéodentines montrent de grandes analogies avec l’os dans l’orientation spatiale de leurs composants («ostéones de dentine» des dents de divers Chondrichthyens). À côté de la dentine proprement dite, on range des tissus dentineux variés (mésodentine, semidentine), connus chez certains Ostéostracés (Agnathes), Acanthodiens et Arthrodires (Placodermes).
Le cément , tissu paradentaire, ressemble beaucoup à l’os tant par sa structure que par sa composition chimique. Chez les Mammifères, il faut distinguer le cément de la couronne dentaire (Rongeurs, Ruminants, chevaux, etc.) du cément radiculaire. Le cément coronal contient des cellules (cémentocytes) et il est modérément minéralisé; le cément radiculaire n’en contient pas toujours. Le premier serait particulier aux Mammifères; le cément radiculaire, par contre, paraît répandu chez tous les Vertébrés.
2. Anatomie comparée
Le type morphologique le plus simple est la dent conique à une pointe, ou dent unicuspidée. Caractéristique de beaucoup de Vertébrés inférieurs, ce type, dit haplodonte (du grec haploos : simple), correspond pour la dent à une fonction de préhension ou de rétention. À ce type s’oppose la dent plexodonte , dent compliquée, à plusieurs tubercules ou cuspides. Caractéristique de la dent jugale des Mammifères, elle revêt une infinité de formes correspondant aux différentes modalités de la fonction de mastication.
Chez un animal donné, les dents peuvent être toutes semblables, (homodontie ), cas fréquent des dents haplodontes des Vertébrés inférieurs. Si la série dentaire montre au contraire des dents dissemblables, il y a hétérodontie ; c’est la règle générale chez les Mammifères.
Mode d’insertion des dents
Les dents des Chondrichthyens sont fixées aux mâchoires par une gaine de derme dense fibreux qui enserre leur plaque basale. Ce type d’insertion autorise des déplacements latéraux importants des dents, ce qui facilitera leur remplacement continuel.
Chez certains Téléostéens (Lophius , Merlucius ), les dents sont attachées à la mâchoire par un ligament très élastique («dents à ressort»): la dent s’efface devant la proie puis se redresse, empêchant toute fuite (fig. 4). Chez beaucoup de Poissons, Amphibiens et Reptiles, il y a au contraire attache par ankylose. La base de la dent se soude, par l’intermédiaire d’un socle calcifié (cément radiculaire?), à la surface des mâchoires. Si la dent se soude par sa base comme chez certains lézards (Agamidés), on parle d’attache de type acrodonte . Lorsqu’elle se soude latéralement comme chez d’autres lézards, les Iguanes, l’insertion est dite pleurodonte . Enfin, la dent peut s’insérer par enchâssement (insertion thécodonte ) dans un alvéole du squelette de la mâchoire (fig. 5). Cette disposition se rencontre chez quelques Poissons (Lepidosteus , Sargus ), chez les nombreux Reptiles Archosauriens fossiles (Thécodontes, Dinosaures) ou actuels (crocodiles), chez les Mammifères enfin.
D’un point de vue fonctionnel, la dent ne peut être dissociée de son appareil d’insertion (appareil péridentaire), qui joue notamment un rôle amortisseur très important.
Mode de remplacement dentaire
On distingue deux modalités de remplacement dentaire: le remplacement latéral et le remplacement vertical. Dans le remplacement latéral , une série de dents de remplacement s’échelonne sur le côté lingual de chaque dent fonctionnelle, elles sont couchées ou déjà redressées, selon les espèces et forment avec la dent en fonction une «famille dentaire». Quand la dent en fonction tombe, la première dent de remplacement qui la côtoie immédiatement vient occuper sa place, et ainsi de suite. Cette modalité de remplacement, très fréquente, se rencontre chez les Chondrichthyens, de nombreux Poissons osseux et divers Reptiles actuels et fossiles. Dans le mode de remplacement vertical , le germe de la dent de remplacement se forme apparemment sous la dent fonctionnelle (majorité des Mammifères; quelques Poissons, tel Pseudoscarus ; Oiseaux fossiles dentés, tel Hesperornis ). Pourtant, chez des Vertébrés très primitifs, les Ostracodermes, les odontodes d’une nouvelle génération se forment dans la peau au-dessus des odontodes de la génération immédiatement antérieure: c’est apparemment l’inverse de la situation observée chez les Mammifères. L’observation embryologique du germe dentaire mammalien viendrait lever cette difficulté: au début du développement, le germe de la dent permanente est placé à côté et au-dessus de celui de la dent de lait; conservant la même relation topographique que chez les Ostracodermes, sa migration sous la dent de lait n’interviendra qu’ultérieurement. Les crocodiles et Anoures ont d’ailleurs un mode de remplacement pseudovertical: la dent de remplacement se différencie d’abord à côté de la dent fonctionnelle et ne vient se placer que secondairement sous elle. Chez l’éléphant et le lamantin, le remplacement, très particulier, s’effectue horizontalement, d’arrière en avant. On voit donc que les remplacements vertical et latéral ne sont pas des modalités fondamentalement irréductibles mais sont réunis par des intermédiaires.
Dents et squelette dermique
Les Agnathes anciens (Ostracodermes) étaient fréquemment revêtus d’un exosquelette très développé, dont la structure histologique a été minutieusement étudiée (Ørvig, Tarlo). La partie externe de cet exosquelette était constituée de tubercules coniques (odontodes) disposés régulièrement à la limite dermo-épidermique. Chaque odontode était formé par un cône de dentine; seule la pointe de l’odontode devait percer l’épiderme. La structure des odontodes est donc absolument comparable à celle de dents qui seraient toutes soudées sur une base osseuse commune. Dans certains cas au moins, la structure fine de la dentine paraît déjà identique à celle de l’homme. Ainsi, un caractère intéressant de ces premiers Vertébrés encore démunis de mâchoire est la présence fréquente dans leur exosquelette d’odontodes qui seraient les précurseurs phylogéniques des dents. Celles-ci ne correspondraient qu’à une différenciation, au niveau buccal, de telles structures exosquelettiques cutanées, après que la modification des arcs branchiaux antérieurs ait permis l’acquisition d’une mâchoire (Gnathostomes).
En effet, d’après les travaux d’Hertwig (1874), on admet universellement que les dents des Chondrichthyens sont homologues aux écailles placoïdes qui recouvrent tout le corps de ces animaux; elles ne constitueraient donc qu’une différenciation plus poussée de ces écailles au niveau buccal. L’écaille placoïde, très évoluée, correspondrait à un synchronomorium (cf. infra , théorie lépidomoriale). D’autre part, les écailles placoïdes des Chondrichthyens et les dents des Vertébrés tétrapodes seraient des formations homologues (Hertwig). Dès lors, les dents apparaissent comme homologues à des éléments du squelette dermique des Vertébrés inférieurs; ce sont des constituants confinés à la cavité buccale des Vertébrés supérieurs.
Écailles et dents sont encore pratiquement identiques chez les Chondrichthyens, mais ces deux types d’organes ne tardent pas à diverger considérablement les uns par rapport aux autres dans les autres groupes de Vertébrés inférieurs, soit pour constituer plaques et écailles, soit pour se spécialiser au niveau buccal sous forme de dents. Chez tous les Poissons osseux, les tissus dentaires de l’exosquelette tendent en général à se réduire des formes primitives aux formes modernes. Cela est vrai pour de nombreuses lignées indépendantes et ce processus constitue un excellent exemple d’évolution parallèle portant sur la régression de ces éléments du squelette.
Chez les Vertébrés tétrapodes, les tissus dentaires disparaissent complètement du squelette dermique.
Les Amphibiens fossiles possèdent souvent des dents à replis compliqués de dentine (plicidentine), d’où le nom de Labyrinthodontes qui les désigne fréquemment. La similitude de structure avec la dent des Crossoptérygiens est un argument parmi beaucoup d’autres pour faire provenir les tétrapodes de ce groupe de Poissons. La plicidentine des Labyrinthodontes était recouverte d’une substance adamantoïde dont la nature exacte est inconnue. Chez les Amphibiens modernes, à dents petites et faibles, une très mince couche d’un tissu adamantoïde ectodermique recouvrirait une différenciation adamantoïde mésodermique périphérique de la dentine. La dent des Reptiles comprend de l’orthodentine, parfois de la plicidentine (varans). La dentine est recouverte par un émail vrai (ectodermique), mais non disposé en «prismes» comme celui des Mammifères. On connaît toutefois des «pseudo-prismes» d’émail chez certains Reptiles fossiles. Chez ceux-ci, comme chez l’alligator actuel, l’émail comporte en fait deux couches, l’une profonde, prismatique, dissimulée par une couche superficielle aprismatique.
Enfin, les Mammifères ont des dents fondamentalement constituées d’orthodentine et d’émail prismatique. Le cément coronal est abondant chez les Artiodactyles, Périssodactyles, Proboscidiens et Rongeurs. Chez les éléphants, la dentine des défenses, ou ivoire proprement dit, a une structure particulière, bien visible sur une section transversale de défense et donnant aux objets en ivoire leur aspect caractéristique.
Chez les Mammifères comme chez les autres Vertébrés, une dentine «secondaire» est susceptible de se former à la périphérie de la cavité pulpaire, mais le processus est assez lent et limité chez l’homme. Contrairement à une opinion très répandue, les dents humaines seraient donc néanmoins capables d’une certaine autoréparation.
Morphologie de la denture
Poissons
Les Placodermes n’ont pas de véritables dents, mais plutôt des plaques dentaires assez particulières. Chez les requins, les dents sont de formes assez variées, dérivées du type haplodonte par un aplatissement plus ou moins prononcé. Elles portent parfois des crénelures périphériques. Certaines raies ont des dents «en pavés» (Myliobates broyeurs de coquillages).
Les Poissons osseux auraient généralement des dents typiquement haplodontes, coniques, nombreuses (polyodontie) et toutes semblables (homodontie). Cette assertion ne doit pas faire illusion: on est accoutumé à croire que seuls les Mammifères possèdent une denture franchement hétérodonte, avec différenciation en incisives, canines et molaires; en fait, l’hétérodontie est relativement répandue dans les diverses espèces de Vertébrés. Les daurades, qui consomment des coquillages, ont des dents broyeuses molariformes à couronne aplatie. Au total, les Poissons possèdent des dents de morphologie étonnamment variée. Chez ces animaux, les dents peuvent se former à la surface de nombreux os dermiques de la région bucco-pharyngée. Cette faculté, particulière aux Poissons, de différenciation des dents sur un territoire encore très vaste constitue un caractère primitif: chez les Vertébrés tétrapodes, la répartition des dents ira progressivement en se réduisant.
Amphibiens
Les Labyrinthodontes avaient de grandes dents coniques à hétérodontie relativement peu accentuée; elles se répartissaient sur les bords des mâchoires, mais aussi sur de nombreux os du palais. Chez les Anoures, Urodèles et Apodes actuels, les dents sont réduites (parfois absentes), coniques, pédicellées, à extrémité souvent bicuspide. Les Anoures ont encore des dents vomériennes sur le palais.
Reptiles
Il y a en général polyodontie et homodontie; les dents sont de type haplodonte cylindro-coniques comme chez les lézards et les crocodiles, ou incurvées vers l’arrière comme chez beaucoup de serpents. Les Reptiles fossiles avaient des dents de morphologie plus variée que les formes actuelles. Les Placodontes, Sauroptérygiens marins, avaient quelques énormes dents en pavés: c’étaient des broyeurs de coquillages. Certains Dinosaures ornithopodes (Trachodon ) avaient de petites dents broyeuses très nombreuses et assez différenciées, serrées les unes contre les autres, et formant ainsi de vastes batteries d’abrasion: c’est le type d’adaptation des énormes dents des Proboscidiens qui est réalisé là, au moyen d’un dispositif anatomique fondamentalement différent (analogie fonctionnelle). Les Reptiles mammaliens (Thérapsidés), précurseurs des Mammifères, présentent souvent une hétérodontie prononcée, avec incisives, canines et postcanines différenciées. Les postcanines étaient déjà des dents pluricuspidées dans certains cas (Cynodontes, Tritylodontes). Ces transformations seraient associées au développement du palais secondaire et à l’apparition de la mastication.
Les serpents montrent une intéressante évolution morphologique des dents, liée à un perfectionnement de la fonction venimeuse.
Mammifères
Les dents sont confinées aux os prémaxillaire, maxillaire et dentaire. L’oligodontie et l’hétérodontie sont la règle. On distingue en avant les incisives préhensiles et coupantes, souvent à une seule racine, et les canines, lacératrices, coniques, monoradiculées. Les dents jugales sont en avant les prémolaires et en arrière les molaires, multicuspidées (plexodontie ) et multiradiculées. Le nombre des dents, en général constant dans une espèce donnée, permet d’écrire la formule dentaire qui tient toujours compte d’une hémimâchoire. Pour l’homme, par exemple, la formule dentaire s’écrira:
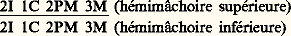
On définira simplement quelques types d’adaptation de la denture mammalienne (fig. 6, 7, 8) .
L’adaptation au régime carné se caractérise par des incisives moyennement ou peu développées, mais les canines sont très grandes (crocs). Les dents jugales tranchantes correspondent au type sécodonte . Dans le cas des Fissipèdes (chien, par exemple) les plus grosses dents, ou carnassières, coupent la nourriture grâce aux mouvements verticaux, en coup de ciseau, de la mandibule, dont la largeur est inférieure à celle de la machoire supérieure, ce qui permet le recouvrement des carnassières du bas par celles du haut lors de l’occlusion maxillaire.
L’adaptation au régime végétarien, dont le type se rencontre chez les Ongulés, s’accompagne d’un diastème (interruption de la série dentaire) entre les incisives-canines et les prémolaires-molaires. Les dents antérieures régressent ou disparaissent souvent, tandis qu’il y a fréquemment molarisation des prémolaires qui forment avec les molaires une batterie dentaire jugale homogène, constituant une table d’abrasion. Dans les formes primitives ou peu spécialisées (régime omnivore), les tubercules dentaires restent bas, hémisphériques: type bunodonte (Suidés). Dans les formes plus évoluées, les dents jugales possèdent des crêtes «en croissant de lune»: type sélénodonte (Ruminants), lorsque les mouvements de broyage se font dans le sens transversal, comme chez les Bovidés.
Si les mouvements de broyage sont antéro-postérieurs, les crêtes des dents jugales sont transversales: type tœchodonte (éléphants, Rongeurs). Dans la lignée du cheval, la dentition jugale est passée d’un type bunodonte (Hyracotherium de l’Éocène) à un type sélénodonte (Merychippus du Miocène), puis lophodonte chez Equus , où des crêtes secondaires donnent à la dent un relief caractéristique, tandis que la couronne dentaire devient de plus en plus haute.
D’autres Périssodactyles (rhinocéros, tapirs) ont conservé au contraire une denture beaucoup plus primitive, avec une couronne basse, à dessin tœchodonte (tapir).
D’une façon très générale, les omnivores tendent à conserver une denture relativement complète, peu spécialisée, sans diastèmes. On peut rattacher à cette tendance la denture des Insectivores, Chiroptères et Primates, dont les dents jugales ont une couronne basse.
En conclusion, il importe d’insister sur l’exceptionnel intérêt que présentent les dents des Mammifères pour les études de taxinomie et d’évolution. La morphologie de la dent mammalienne exprime de façon subtile et précise les adaptations variées du régime alimentaire et constitue un excellent caractère taxinomique permettant de différencier les formes animales jusqu’au niveau de l’espèce (c’est-à-dire deux espèces voisines).
3. Quelques problèmes généraux
Origine des dents jugales des Mammifères
Ce problème, qui est celui de l’origine de la plexodontie, a suscité de nombreuses théories.
Les théories «explicatives » s’appuient essentiellement sur l’anatomie comparée et sur l’embryologie. On a ainsi supposé que les nouvelles cuspides seraient apparues par «bourgeonnement», par «irritation fonctionnelle», par «concrescence», par «fusion de germes dentaires», soit de même génération, soit de générations successives (Bolk). Dans l’état actuel des connaissances, aucune de ces conceptions ne semble plus capable de rallier les suffrages.
Les théories «phylogénétiques » tentent de reconstituer l’évolution de la plexodontie en s’appuyant sur la paléontologie.
La théorie de la multituberculie , développée par M. Friant et R. Anthony (1932), propose de voir dans les molaires multicuspidées des Multituberculés un type (bunodonte) tout à fait généralisé, correspondant à un régime omnivore, et qui serait à l’origine des autres types de dents jugales de tous les Mammifères. Cette conception ne semble pas avoir résisté à l’épreuve du temps.
La théorie lépidomoriale de Stensiö et Ørvig (1961) a été d’abord développée pour rendre compte de l’origine des écailles placoïdes des Sélaciens. Stensiö et Ørvig ont montré que l’écaille placoïde, bien loin de constituer une unité structurale primitive, était au contraire une formation complexe, résultat d’un très long processus évolutif. Les éléments primordiaux de l’exosquelette seraient de minuscules cônes de dentine, les lepidomoria , différenciés à la limite dermo-épidermique au-dessus d’une anse vasculaire. Ces lepidomoria pourraient fusionner entre eux précocement pour former des écailles synchronomoriales plus complexes. Les synchronomoria à leur tour s’assembleraient en écailles cyclomoriales plus grandes, à partir desquelles on pourrait faire dériver l’écaille placoïde.
Comme la dent conique des premiers tétrapodes paraît clairement dérivée d’un type placoïde, elle serait donc passée au cours de son évolution première par les étapes successives décrites ci-dessus: sa simplicité ne serait qu’apparente. Dans cette perspective, l’apparition de la dent pluricuspidée des Mammifères ne serait qu’une manifestation (tardive) des potentialités morphogénétiques variées que la dent haplodonte (écaille placoïde) aurait héritées de son histoire antérieure complexe. Si la théorie lépidomoriale, basée sur des observations irréprochables, paraît extrêmement féconde pour l’étude des tissus durs des Vertébrés inférieurs, beaucoup pensent que l’application que l’on veut en faire pour rendre compte de la plexodontie des Mammifères est loin d’être pleinement convaincante.
Le néotrituberculisme , universellement accepté par les chercheurs, rend compte des faits connus et a apporté aussi une précieuse terminologie (fig. 6 et 7) . Dû à Cope et Osborn (1874), modifié et complété par Gregory (1910) et Simpson (1936), le trituberculisme pose essentiellement le principe d’une différenciation progressive des dents jugales mammaliennes et se propose de mettre en évidence les homologies entre les diverses cuspides dentaires. Selon ces conceptions, il faudrait distinguer un stade primitif (trigonodonte ), ou «trigone primordial», comprenant trois cuspides. Ce stade évolutif primitif est représenté par les molaires du Mammifère jurassique Dryolestes . Ultérieurement, un «stade tribosphénique» se serait différencié: il est atteint chez le Mammifère paléocène Deltatheridium . Toutes les dents jugales mammaliennes pourraient dériver de ce schéma fondamental.
Si, en effet, chez les premiers Mammifères (non thériens), les cuspides des molaires sont alignées et non disposées en triangle (Triconodontes, Docodontes, Haramyidés), les molaires des Kuehneothériidés du Rhétien et des Symmétrodontidés du Jurassique et du Crétacé inférieur montrent trois cuspides disposées en triangle. Un stade intermédiaire entre cette structure trigonodonte et la molaire tribosphérique des Thériens (chez lesquels les cuspides sont plus nombreuses) pourrait être représenté par la structure observée chez les Eupantothériens (Jurassique-Crétacé inférieur). Dans ce groupe, en effet, si le protocône est absent sur les molaires supérieures, un talonide s’ajoute au trigonide sur les molaires inférieures.
Les découvertes paléontologiques ont mis en évidence une complication progressive et ménagée de la denture à travers toute la série Synapside. Très progressivement et simultanément, le passage se fait de l’homodontie à l’hétérodontie, de la polyodontie à l’oligodontie, de l’haplodontie à la plexodontie. Ainsi, dans une large mesure, les documents actuels réduisent-ils le hiatus morphologique entre dent reptilienne et dent mammalienne, qui avait frappé les théoriciens d’il y a quelques décennies.
Nombre de générations dentaires
Les polyphyodontes possèdent un grand nombre de dentitions successives, toutes semblables; ce sont les Poissons (jusqu’à cent dentitions successives), les Reptiles (crocodiles et serpents: jusqu’à vingt-cinq dentitions successives). Les oligophyodontes sont essentiellement les Mammifères: le nombre de générations dentaires est réduit et les dentitions qui se succèdent ne sont pas semblables. Les diphyodontes sont les oligophyodontes à deux dentitions, une lactéale et une permanente (majorité des Mammifères actuels, tel l’homme). Les monophyodontes n’ont qu’une seule dentition. Parmi eux, les Édentés et quelques Chiroptères (chauve-souris) ont quand même une dentition lactéale qui régresse tôt et n’est jamais fonctionnelle (monophyodontie apparente). Au contraire, chez les Cétacés odontocètes (dauphins) et les Proboscidiens, seule la dentition lactéale est présente et fonctionnelle toute la vie. La dentition définitive n’existe jamais.
Chez les diphyodontes, certaines dents appartiennent à une dentition unique et ne sont pas remplacées: ce sont les dents monophysaires (ex.: les molaires de l’homme). De nombreux arguments morphologiques et paléontologiques permettent d’interpréter ces dents comme des dents lactéales qui auraient seulement subi un retard dans leur développement et leur éruption: ainsi, par exemple, les molaires humaines définitives auraient plus d’affinités morphologiques avec les molaires de lait qu’avec les prémolaires définitives.
Modes de croissance dentaire
Chez les polyphyodontes, la croissance de chaque dent est limitée et les dents sont remplacées au cours de toute l’existence. Chez les oligophyodontes, les dents peuvent croître selon trois modalités (fig. 8). Dans un premier cas, la croissance est limitée dans le temps, la couronne dentaire est basse (type brachyodonte ), la racine est allongée et se ferme assez rapidement à sa base (Insectivores, Carnassiers, Primates). Chez divers herbivores, les dents jugales subissent une forte abrasion mécanique de la couronne, ceci est compensé par une croissance de la dent prolongée une partie de la vie, la racine est courte et reste ouverte (type hypsodonte ) et la couronne est très haute (Ruminants, chevaux, éléphants, certains Rongeurs comme le porc-épic). Enfin, certaines dents à usure très rapide se développent toute la vie, la racine est réduite, ouverte (incisives de l’éléphant, des Rongeurs, «corne» du narval, canines des porcs, molaires de certains Rongeurs comme le castor, d’Édentés comme le paresseux ou de l’oryctérope, un Tubulidenté). Les modalités de croissance des dents apparaissent comme des caractères adaptatifs, sans doute aussi importants que l’évolution du dessin des crêtes coronales. En général, dans une lignée évolutive donnée, il y a passage d’un état brachyodonte primitif à un état hypsodonte évolué (lignée du cheval), sans que cela soit nécessairement réalisé selon une modalité «orthogénétique». L’acquisition d’une croissance dentaire continue paraît être une nécessité mécanique impérative dans le cas de certains régimes alimentaires herbivores, dès lors que l’oligophyodontie remplace la polyphyodontie.
Régression dentaire. Anodontie
Dans certaines lignées appartenant à toutes les classes de Vertébrés, les tissus dentaires régressent ou même disparaissent complètement. Chez les lamproies et les myxines, des formations odontoïdes, d’origine uniquement épidermique et de nature cornée, sont disposées sur les parois de la gueule et sur la langue. Des Poissons comme l’esturgeon et l’hippocampe sont entièrement dépourvus de dents; chez les Amphibiens, il en va de même des crapauds. Des Reptiles comme les tortues présentent un bec corné et sont totalement privés de dents, contrairement à leurs ancêtres du Trias. Chez les Oiseaux, on connaît l’existence de dents chez l’Archaeopteryx du Jurassique supérieur et chez les formes encore primitives de la fin du Secondaire (Ichthyornis et Hesperornis ); les formes actuelles sont entièrement anodontes. Selon des expériences embryologiques, les cellules aviennes peuvent induire la différenciation de tissus dentaires: les gènes correspondants n’auraient donc pas été «perdus» au cours de l’évolution des Oiseaux post-crétacés, mais resteraient muets depuis 60 millions d’années! Parmi les Mammifères, l’échidné dépourvu de dents présente un bec corné; l’ornithorynque est aussi anodonte au stade adulte. Les Édentés et les Pholidotes ont des dents régressées par quasi-disparition ou disparition totale de l’émail; les fourmiliers et les pangolins en manquent totalement. Les Cétacés présentent une intéressante évolution régressive des dents. Les formes primitives, comme Pappocetus de l’Éocène, étaient des diphyodontes hétérodontes, condition habituelle des Mammifères. Les Odontocètes actuels (dauphins, marsouins) sont revenus à la polyodontie (200 dents pour le dauphin) et à l’homodontie haplodonte (dents simples toutes semblables), situation fréquente chez les Reptiles et les Poissons. Toutefois, ces Cétacés sont monophyodontes; le cachalot n’a que 40 à 50 dents et seulement à la mandibule, le narval n’a que deux incisives, la gauche du mâle formant l’énorme «dent de licorne» qui peut atteindre deux mètres. Enfin, les Cétacés à fanons ou mysticètes (baleine) sont anodontes, bien que des germes dentaires se développent avant la naissance, mais ils ne font jamais éruption. Les fanons sont des productions kératinisées issues de l’épiderme de la mâchoire supérieure: ils jouent un rôle de filtre et constituent donc une adaptation au régime planctonophage.
Ainsi, la tendance à la disparition des tissus dentaires se manifeste indépendamment dans certaines lignées, sans qu’il soit toujours possible de rattacher cette évolution à une adaptation nette à l’égard d’un régime alimentaire précis. Les organes cornés, qui jouent très souvent chez les anodontes un rôle analogue à celui de la denture et semblent venir, en quelque sorte, «compenser» sa perte, varient beaucoup dans les différents groupes, souvent en fonction des adaptations particulières, mais ils peuvent aussi fournir parfois d’excellents exemples de convergence morphologique, comme les becs de l’ornithorynque et du canard.
Évolution et signification des tissus dentaires
On a cru longtemps que le cartilage avait précédé tous les autres tissus durs au cours de l’évolution des Vertébrés: l’os, la dentine, l’émail seraient des tissus plus évolués et apparus ultérieurement. La paléontologie moderne s’inscrit en faux contre cette opinion, issue des idées haeckeliennes. Actuellement, on pense que tous les tissus durs mésodermiques sont d’ancienneté égale. Chez les Vertébrés les plus anciens, la présence simultanée de tous les tissus durs mésodermiques, ainsi qu’une certaine interpénétration de leurs caractéristiques (ce qui rend leur classification si difficile), semble indiquer qu’ils ont tous entre eux des rapports étroits; au cours du déroulement ultérieur de l’évolution, on assisterait à une certaine divergence de ces tissus, qui atteignent chacun leur pleine identité, telle qu’on peut l’observer chez les Mammifères. Chacun de ces tissus (cartilage, os, dentine, cément) paraît alors adapté à un rôle bien particulier, mais cette apparente spécialisation ne doit pas faire illusion: dans une certaine mesure, les dents offrent un exemple de l’«opportunisme de l’évolution» (Simpson) qui a utilisé à une fonction particulière les tissus durs du squelette dermique. Certaines caractéristiques troublantes et très «actuelles» des dents humaines, telle l’hypersensibilité de la dentine mise à nu, ne sont peut-être que le témoignage de la fonction primitivement sensorielle de ce tissu (Tarlo). Les tissus dentaires ne sont certainement pas apparus chez les Vertébrés uniquement «pour» remplir les seules fonctions spéciales de préhension et de concassage des aliments, puisque toutes ces fonctions peuvent très bien s’accomplir en leur absence, comme le montrent les lignées anodontes.
Encyclopédie Universelle. 2012.