процесс, заключающийся а серии клеточных дифференцировок, в результате которых образуются зрелые клетки крови.
Во взрослом организме существуют родоначальные кроветворные, или стволовые, клетки. Предполагают, что они закладываются в эмбриогенезе в относительно небольшом количестве (порядка сотен тысяч — миллионов). По мере необходимости эти клетки одна за другой вступают в дифференцировку, образуя категорию более дифференцированных кроветворных клеток. Стволовые кроветворные клетки в стадии созревания находятся под строгим регулирующим контролем, механизм которого изучен не полностью. На ранних этапах созревания важное значение имеют, по-видимому, локальные факторы, продуцируемые стромальными клетками, т.е. кроветворным микроокружением. Влияние микроокружения осуществляется путем взаимодействия стромальных и кроветворных (в первую очередь стволовых) клеток. Такие регуляторные взаимодействия требуют прямых клеточных контактов. При этом образуются своеобразные структуры — клеточные островки, представляющие собой группы кроветворных клеток, которые лежат в сети отростков ретикулярных клеток, адвентициальных клеток синусов костного мозга. Механизм функционирования таких островков не известен. Возможно в нем принимают участие местнодействующие гормоны, происходит непосредственное взаимодействие клеточных поверхностей или формируются трансмембранные переходы. В регуляции К. принимают участие также цитокины (итерлейкины, колониестимулирующий фактор, факторы роста), гормоны (бурстопромоторная активность) и другие гуморальные факторы, например гемопоэтины, к которым относят эритропоэтины, лейкопоэтины, тромбопоэтины.
Дифференцировка стволовой кроветворной клетки в первые морфологически распознаваемые клетки того или иного ряда представляет собой многостадийный процесс, ведущий к значительному увеличению численности каждого из рядов (рис.). На этом пути происходит постепенное ограничение способности клеток-предшественниц (этим термином обозначают всю совокупность морфологически сходных клеток верхних трех рядов схемы кроветворения) к различным дифференцировкам и постепенное снижение их пролиферативного потенциала.
В дальнейшем удалось обнаружить поэтино-чувствительные клетки-предшественницы. Абсолютное большинство их находится в стадии активной пролиферации. Морфологически они, так же как и стволовые клетки, не отличимы от лимфоцитов. Принципиальной особенностью поэтинчувствительных клеток является их способность отвечать на гуморальные регулирующие воздействия. Между стволовыми и поэтиночувствительными клетками находятся клетки-предшественницы миелопоэза и лимфоцитопоэза. Существование этих клеток строго не доказано, однако установлено, что при ряде лейкозов, прежде всего хроническом миелолейкозе, сублейкемическом миелозе, эритромиелозе (см. Лейкозы), единственным источником опухолевой пролиферации могут быть клетки более молодые (менее дифференцированные), чем поэтиночувствительные, но более зрелые, чем стволовые. Выявлены также лимфолейкозы, характеризующиеся одновременным поражением В- и Т-лимфоцитов, т. е. возникшие из их общего предшественника.
В схеме К. стволовая клетка и клетки второго и третьего рядов взяты в рамки и даны в двух морфологически разных вариантах, в которых они способны находиться: лимфоцитоподобном и бластном (бластом называется клетка, имеющая обычно неширокую цитоплазму, нежноструктурное ядро, отличающееся равномерностью калибра и окраски хроматиновых нитей, часто содержащее нуклеолы).
На уровне поэтинчувствительных клеток происходит дальнейшее ограничение дифференцировочных возможностей клеток. На этой и следующих морфологически распознаваемых стадиях дифференцировки подавляющее большинство клеток находится в состоянии пролиферации.
Последними клетками, способными к делению среди гранулоцитов являются миелоциты, а среди эритрокариоцитов — полихроматофильные нормоциты. В процессе дифференцировки морфологически распознаваемые клетки эритроцитарного ряда претерпевают 5—6 митозов; гранулоцитарные клетки — 4 митоза; при моноцитопоэзе от монобласта до макрофага происходит 7—8 митозов. В мегакариоцитопоэзе выделяют несколько морфологически различимых предшественников, которые начиная с мегакариобласта претерпевают 4—5 эндомитозов (деления ядра без деления цитоплазмы).
С помощью метода клонирования и анализа хромосомных маркеров было показано, что фагоцитирующие клетки, в частности купферовские клетки печени и все другие тканевые макрофаги, объединенные в систему фагоцитирующих мононуклеаров, относятся к производным кроветворных клеток и представляют собой потомство моноцитов, а не ретикулярных и эндотелиальных клеток. Клетки этой системы не имеют гистогенетической общности ни с ретикулярными, ни с эндотелиальными клетками. Основные функциональные характеристики, присущие входящим в эту систему клеткам, включают способность к фагоцитозу, пиноцитозу, прочному прилипанию к стеклу. По мере дифференцировки в клетках этого ряда появляются рецепторы для иммуноглобулинов и комплемента, благодаря чему клетки приобретают способность к активному фагоцитозу.
В эритроцитопоэзе самой молодой клеткой является эритробласт (ее называют также проэритробластом), который имеет бластную структуру и обычно круглое ядро. Цитоплазма при окраске темно-синяя, располагается узким ободком, часто образует своеобразные выросты. В отношении клеток эритрокариоцитарного ряда нет единой номенклатуры. Их называют как нормобластами, так и эритробластами. Поскольку для других рядов термин «бласт» применяется лишь для клеток-родоначальниц того или иного ростка (отсюда и название «бласт» — росток), все клетки, являющиеся потомством эритробласта, должны иметь в названии окончание «цит». Поэтому термин «нормобласты» был заменен на «нормоциты».
За эритробластом появляется пронормоцит, который отличается от эритробласта более грубым строением ядра, хотя оно и сохраняет правильную структуру хроматиновых нитей. Диаметр ядра меньше, чем у эритробласта, ободок цитоплазмы шире, видна перинуклеарная зона просветления. При изучении миелограммы пронормоцит легко спутать с эритробластом. В связи с трудностью разделения этих клеток некоторые авторы предлагают в практической гематологии их вообще не дифференцировать. Далее следует базофильный нормоцит, у которого грубоглыбчатое ядро имеет колосовидную структуру, а цитоплазма окрашена в темно-синий цвет. Следующий — полихроматофильный нормоцит отличается еще более плотной структурой ядра; цитоплазма занимает бо́льшую часть клетки и имеет базофильную (за счет структур, содержащих РНК) и оксифильную (в связи с появлением достаточного количества гемоглобина) окраску.
Оксифильный или ортохромный нормоцит содержит маленькое плотное ядро как вишневая косточка), оксифильную или с базофильным оттенком цитоплазму. В норме оксифильных нормоцитов сравнительно мало, т.к., выталкивая на этой стадии ядро, клетка превращается в эритроцит, но в «новорожденном» эритроците всегда сохраняются остатки базофилии за счет небольшого количества РНК, которая исчезает в течение первых суток. Такой эритроцит с остатками базофилии называется полихроматофильным эритроцитом. При применении специальной прижизненной окраски базофильное вещество выявляется в виде сеточки; тогда эту клетку называют ретикулоцитом.
Зрелый эритроцит имеет форму двояковогнутого диска, поэтому в мазке крови он имеет центральное просветление. По мере старения форма эритроцита постепенно приближается к сферической.
Самой молодой клеткой тромбоцитопоэза является мегакариобласт — одноядерная небольшая клетка с крупным бластным ядром, хроматинные нити которого толще и грубее, чем у эритробласта; в ядре могут быть видны 1—2 темно-синие нуклеолы. Цитоплазма беззернистая, темно-синего цвета, отростчатая, узким ободком окружает ядро. Промегакариоцит возникает в результате нескольких эндомитозов. Ядро полиморфное, с грубым строением хроматина; цитоплазма темно-синяя, беззернистая.
Зрелый мегакариоцит отличается от промегакариоцита большим ядром. Цитоплазма имеет сине-розовую окраску, содержит азурофильную красноватую зернистость. Внутри мегакариоцита формируются тромбоциты. В мазке крови можно видеть и распадающиеся мегакариоциты, окруженные кучками тромбоцитов. При тромбоцитолитических состояниях отшнуровка тромбоцитов может происходить и на стадии промегакариоцита; тромбоциты при этом лишены азурофильной субстанции, но активно участвуют в Гемостазе.
Лейкоцитопоэз включает гранулоцитопоэз, лимфоцитопоэз и моноцитопоэз. В гранулоцитарном ряду миелобласт представляет первую морфологически различимую клетку. Он имеет нежноструктурное ядро, единичные нуклеолы. Форма ядра круглая, размеры чуть меньше, чем у эритробласта. Миелобласт отличается от недифференцируемых бластов из класса клеток-предшественниц наличием зернистости в цитоплазме; форма клетки чаще круглая, ровная.
На следующей стадии гранулоцитопоэза обнаруживается промиелоцит — нейтрофильный, эозинофильный и базофильный. Круглое или бобовидное ядро промиелоцита больше ядра миелобласта почти вдвое, хотя эта клетка и не является полиплоидией; оно часто располагается эксцентрично, и в нем можно видеть остатки нуклеол. Структура хроматина уже утрачивает нежное нитчатое строение бластных клеток, хотя и не приобретает грубоглыбчатого строения. Площадь цитоплазмы примерно равна площади ядра; цитоплазма обильно насыщена зернистостью, имеющей характерные для каждого ряда особенности. Для нейтрофильного ряда промиелоцит является самой зернистой клеткой. Его зернистость полиморфная: крупная и мелкая; окрашивается и кислыми, и основными красителями. В промиелоците зернистость часто располагается также на ядре. Зернистость эозинофильного промиелоцита, обладая характерной для эозинофилов однотипностью зерен (типа кетовой икры), вместе с тем окрашивается как кислыми, так и основными красителями. Базофильный промиелоцит имеет крупную полиморфную базофильную зернистость.
Поскольку переход от промиелоцита к миелоциту не является резким, существует промежуточная форма, названная материнским миелоцитом, которая по всем признакам соответствует описанному промиелоциту, но отличается от него более грубым ядром. В практике эта форма не учитывается, в миелограмму она не входит.
Миелоцит представляет собой клетку с круглым или овальным, часто эксцентрически расположенным ядром, потерявшим какие бы то ни было признаки бласта. Цитоплазма окрашена в серовато-синеватый тон, ее зернистость у нейтрофильного миелоцита мельче, чем у промиелоцита. Относительная площадь цитоплазмы увеличивается. Эозинофильный миелоцит имеет характерную однотипную оранжево-красную зернистость, базофильный миелоцит — полиморфную крупную базофильную зернистость.
Метамиелоцит характеризуется бобовидным крупноглыбчатым ядром, расположенным обычно эксцентрично. Площадь его цитоплазмы больше площади ядра; цитоплазма содержит ту же зернистость, что и миелоцит, но в нейтрофильных метамиелоцитах она более скудная, чем в миелоцитах. В палочкоядерных клетках (гранулоцитах) хроматин ядра более плотно упакован, ядро вытягивается, иногда в нем намечается образование сегментов. В зрелых сегментоядерных клетках ядро обычно имеет от 2 до 5 сегментов.
Моноцитарный ряд представлен довольно простыми стадиями перехода. Монобласт в норме трудно отличить от миелобласта или недифференцируемого бласта, но при монобластном остром или моноцитарном хроническом лейкозе эти клетки легко выявить с помощью гистохимической окраски. Промоноцит имеет ядро промиелоцита, но лишен зернистости. Моноциты — наиболее крупные клетки крови; из них происходят макрофаги (см. Система мононуклеарных фагоцитов).
Существенным дополнением к представлению о лимфоцитопоэзе послужило открытие двух типов лимфоцитов — В- и Т-клеток, первые из которых ответственны за гуморальный иммунитет, т. е. выработку антител (Антитела), а вторые обеспечивают клеточный иммунитет, участвуют в реакции отторжения чужеродной ткани (см. Трансплантационный иммунитет). Оказалось, что В-лимфоциты в результате антигенной стимуляции могут из морфологически зрелой клетки превращаться в бластную форму и дальше дифференцироваться в клетки плазматического ряда. Под влиянием антигенной стимуляции трансформируются в бластную форму и Т-лимфоциты. Т.о., ранее казавшийся единым лимфоидный ряд представлен тремя рядами клеток: В-, Т-лимфоцитами и тесно связанными с В-лимфоцитами плазматическими клетками. Кроме того, привычное представление о бластной клетке как о родоначальнице ряда оказалось не совсем точным для лимфоцитов: зрелые лимфоциты при воздействии на них специфических антигенов вновь способны трансформироваться в бластные клетки. Этот феномен получил название реакции бластотрансформации лимфоцитов (см. Лейкоцитарные тесты). Трансформированные под действием антигенов лимфоциты называют иммунобластами. В схему К. пришлось ввести стрелки, указывающие на возможность перехода морфологически зрелых лимфоцитов в соответствующие бластные формы. Лимфобласт (большой лимфоцит) имеет все черты недифференцируемого бласта, но содержит иногда единичные крупные нуклеолы. Обнаружение в мазке из лимфатического узла или селезенки бласта без зернистости позволяет относить его к лимфобластам. Пролимфоцит характеризуется относительно гомогенной структурой ядра, нередко наличием остатков нуклеол, но у него отсутствует характерная для зрелого лимфоцита крупная глыбчатость хроматина.
Плазмобласт имеет бластное ядро, беззернистую фиолетово-синюю цитоплазму. Проплазмоцит по сравнению с плазмоцитом обладает более плотным ядром, расположенным обычно эксцентрично, относительно большей цитоплазмой сине-фиолетового цвета. Плазмоцит отличается колесовидным плотным ядром, лежащим эксцентрично: цитоплазма сине-фиолетовая, иногда с несколькими азурофильными красноватыми гранулами. И в норме, и при патологии он может быть многоядерным.
Будучи гистогенетически единой, кроветворная система в своем функционировании характеризуется определенной независимостью поведения отдельных ростков кроветворения.
Кроветворение осуществляется в кроветворных органах, к которым относят вилочковую железу (Вилочковая железа), Костный мозг, Лимфатические узлы и селезенку (Селезёнка). К. в кроветворных органах, за исключением костного мозга, происходит в основном в антенатальном периоде, а после рождения интенсивность его быстро снижается. Кроветворные органы обладают определенными анатомо-физиологическими особенностями, имеют общие черты строения. Их строму составляет ретикулярная ткань, паренхиму — кроветворные клетки. Эти органы богаты элементами, относящимися к системе мононуклеарных фагоцитов. Характерным является наличие капилляров синусоидного типа. В синусах между эндотелиальными клетками имеются поры, связывающие ткань кроветворных органов с кровяным руслом. Такое строение обеспечивает транспорт клеток крови, а также поступление из крови в кроветворные органы гуморальных факторов (гемопоэтинов), которые наряду с нервной системой оказывают влияние на кроветворение.
Кроветворение в антенатальном периоде впервые обнаруживается у 19-дневного эмбриона в кровяных островках желточного мешка, в стебле и хорионе. К 22-му дню первые кровяные клетки проникают в мезодермальную ткань эмбриона, в сердце, аорту, артерии. На 6-й неделе снижается активность К. в желточном мешке. Полностью первый (мезобластический) период гемопоэза, преимущественно эритроцитопоэза, заканчивается к началу 4-го месяца жизни плода (календарного). Примитивные кроветворные клетки желточного мешка накапливают гемоглобин и превращаются в примитивные эритробласты.
Второй (печеночный) период К. начинается после 6-й недели и достигает максимума к 5-му месяцу. К. этого периода преимущественно эритроидное, хотя на 9-й неделе в печени плода уже созревают первые нейтрофилы. Печеночный период эритроцитопоэза характеризуется исчезновением мегалобластов; при этом эритрокариоциты имеют нормальные размеры. Ни 3-м месяце развития плода в эритроцитопоэз включается селезенка, но у человека ее роль в антенатальном К. ограничена.
На 4—5-м месяце начинается третий (костномозговой) период К. Миелоидный эритроцитопоэз плода относится к эритробластическому и, как и лейкоцитопоэз, мало отличается от эритроцитопоэза взрослого. Общей закономерностью эмбрионального эритроцитопоэза является постепенное уменьшение размеров эритроцитов и увеличение их числа. Соответственно различным периодам К. (мезобластическому, печеночному и костномозговому) существует три разных типа гемоглобина; эмбриональный, фетальный и гемоглобин взрослого. В основном переход от фетального гемоглобина к гемоглобину взрослого начинается на 3-й неделе жизни плода и заканчивается через 6 мес. после рождения.
В первые дни у новорожденных наблюдаются полиглобулия и нейтрофильный лейкоцитоз. Затем активность эритроцитопоэза снижается, нормализуясь в возрасте 2—3 мес. Нейтрофилез первых дней жизни сменяется лимфоцитозом; только к 5 годам в лейкоцитарной формуле начинают преобладать нейтрофилы.
Патология кроветворения может проявляться нарушением созревания клеток, выходом в кровь незрелых клеточных элементов, возникновением в крови несвойственных данному возрасту клеточных элементов. Бактериальная инфекция, обширные тканевые распады (распадающиеся опухоли, флегмоны и др.) сопровождаются выраженным нейтрофильным лейкоцитозом с увеличением процента палочкоядерных нейтрофилов, нередким появлением в крови метамиелоцитов, миелоцитов, промиелоцитов. Четкой зависимости степени лейкоцитоза от тяжести патологического процесса не существует. Лейкоцитоз зависит, с одной стороны, от объема костномозгового и сосудистого гранулоцитарного резерва и от активности костномозговой продукции, с другой — от интенсивности потребления гранулоцитов в очаге воспаления. Противоположное лейкоцитозу состояние — Лейкопения, обусловленное прежде всего гранулоцитопенией, может быть связано с подавлением продукции гранулоцитов в результате воздействия противогранулоцитарных антител, аплазии костного мозга иммунной природы, например характеризующейся одновременным угнетением гранулоцитарного, эритроцитарного и мегакариоцитарного ростков, или аплазии неизвестного происхождения (собственно апластическая анемия); в других случаях гранулоцитопения и лейкопения могут быть вызваны повышенным распадом гранулоцитов в увеличенной селезенке (например, при хроническом гепатите, циррозе печени). В связи с существованием костномозгового резерва уменьшение количества гранулоцитов в крови за счет их повышенного использования встречается редко (например, при обширных сливных пневмониях). Лейкопения является частым признаком опухолевого замещения костного мозга при милиарных метастазах, при острых лейкозах и изредка наблюдается в начале хронического лимфолейкоза. При лейкозах количество лейкоцитов в крови может и увеличиваться; постоянно это происходит при хронических лейкозах. При острых лейкозах содержание лейкоцитов в крови может быть различным: в начале процесса чаще отмечается лейкопения, затем по мере выхода бластных опухолевых клеток в кровь может возникнуть лейкоцитоз.
Вирусная инфекция, антигенные воздействия ведут к усиленной продукции специфических лимфоцитарных клонов, повышению содержания лимфоцитов в крови.
Уменьшение количества тромбоцитов наблюдается при появлении аутоантител к тромбоцитам (реже к мегакариоцитам), при повышенном разрушении их в увеличенной селезенке. Снижение содержания тромбоцитов возможно в результате кровопотерь, при возникновении обширных гематом, внутрисосудистом диссеминированном свертывании (тромбоцитопения потребления). Увеличение содержания тромбоцитов наблюдается при некоторых хронических лейкозах (хроническом миелолейкозе, сублейкемическом миелозе, эритремии), нередко при раке. Иногда при раке почки раковые клетки продуцируют эритропоэтин и, возможно, тромбопоэтин, что сопровождается резким повышением количества эритроцитов и тромбоцитов.
Содержание эритроцитов в крови определяется соотношением их распада и продукции, кровопотерями, обеспеченностью организма железом. Дефицит железа приводит к снижению содержания гемоглобина в эритроцитах при нормальном числе их в крови (низкий цветной показатель). Напротив, дефицит витамина В12 сопровождается нарушением клеточного деления в результате изменений синтеза ДНК; при этом эритроциты уродливы, их мало, но гемоглобина в них больше, чем в норме (повышенный цветной показатель).
В отдельных случаях возможны и реакции нескольких ростков К. на неспецифические стимулирующие воздействия. Так, развитие в организме раковой опухоли может приводить к увеличению в крови содержания как гранулоцитов, так и тромбоцитов. Аналогичная картина изредка наблюдается при сепсисе.
Кроветворение претерпевает глубокие изменения при воздействии ионизирующего излучения (Ионизирующие излучения). Под его влиянием гибнут делящиеся клетки костного мозга, лимфатических узлов. Зрелые гранулоциты, эритроциты сохраняют жизнеспособность даже при заведомо смертельных дозах облучения. Зрелые лимфоциты относятся к радиочувствительным клеткам; этим объясняется быстрое уменьшение их количества в крови в первые же часы после облучения. Поскольку эритроциты в крови живут около 120 дней, анемия развивается через 1—11/2 мес. после облучения. К этому времени в тяжелых случаях начинается активное К., отмечается повышение содержания ретикулоцитов, и анемия не достигает высокой степени. В основном аналогичные изменения развиваются при химиотерапии опухолей.
См. также Костный мозг, Кровь, Лимфатические узлы, Селезенка (Селезёнка).
Библиогр.: Кассирский И.А. и Алексеев Г.А. Клиническая гематология. М., 1970; Максимов А.А. Основы гистологии, ч. 1—2, Л., 1925; Руководство по гематологии, под ред. А.И. Воробьева, М., 1985; Чертков И.Л. и Воробьев А.И. Современная схема кроветворения, Пробл. гематол и перелив. крови, т. 18, №10, с, 3, 1973, библиогр.; Чертков И.Л. и Фриденштейн А.Я. Клеточные основы кроветворения, М., 1977, библиогр.
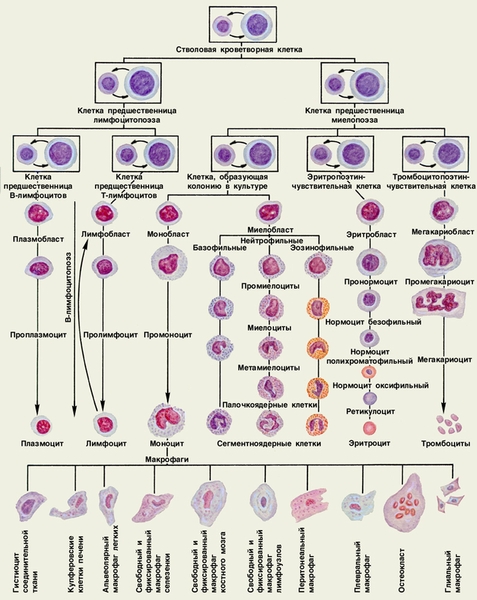
Схема кроветворения. Клетки, изображенные в рамке, даны в двух морфологических вариантах: лимфоцитоподобном (меньшего размера) и бластном (большего размера). Стрелки указывают на возможность перехода клеток одного варианта в клетки другого варианта.
II
Кроветворе́ние (haemopoesis, haemopoiesis; син. гемопоэз)
процесс образования, развития и созревания клеток крови.
Кроветворе́ние ангиобласти́ческое (h. angioblastica) — К. из клеток мезенхимы в желточном мешке и вокруг сосудов в теле зародыша, происходящее одновременно с развитием сосудов.
Кроветворе́ние вика́рное (h. vicaria; син. К. заместительное) — К., характеризующееся заместительной гиперплазией ткани одних кроветворных органов в ответ на снижение функции других.
Кроветворе́ние гетерото́пное (h. heterotopi-са) — К., протекающее в органах и тканях, которые обычно в этом процессе не участвуют.
Кроветворе́ние замести́тельное — см. Кроветворение викарное.
Кроветворе́ние костномозгово́е (h. medullaris) — К. в костном мозге; включает эритро-, гранулоцито- и тромбоцитопоэз.
Кроветворе́ние мегалобласти́ческое (h. megaloblastica) — эритропоэз при Bi2 (фолиево)-дефицитных анемиях, характеризующийся замедлением процесса дифференциации с увеличением количества базофильных эритробластов и наличием морфологически измененных эритробластов — мегалобластов.
Кроветворе́ние нормобласти́ческое (h. normoblastica) — нормальный тип эритропоэза человека в постнатальном периоде, характеризующийся превращением эритробласта в эритроцит через стадии пронормобласта и нормобласта.
Кроветворе́ние экстрамедулля́рное (h. extramedullaris) — К. вне ткани костного мозга (в печени, селезенке, лимфатических узлах и др.), возникающее при некоторых системных болезнях крови.
1. Малая медицинская энциклопедия. — М.: Медицинская энциклопедия. 1991—96 гг. 2. Первая медицинская помощь. — М.: Большая Российская Энциклопедия. 1994 г. 3. Энциклопедический словарь медицинских терминов. — М.: Советская энциклопедия. — 1982—1984 гг.