дезоксирибонуклеиновые и рибонуклеиновые кислоты, универсальные компоненты всех живых организмов, ответственные за хранение, передачу и воспроизведение (реализацию) генетической информации. На два типа все Н. к. делят по углеводному компоненту молекул: дезоксирибозе у дезоксирибонуклеиновых кислот (ДНК) и рибозе у рибонуклеиновых кислот (РНК). Биологическая роль ДНК у большинства организмов заключается в хранении и воспроизведении генетической информации, а РНК — в реализации этой информации в строении молекул белков (Белки) в процессе их синтеза.
Нуклеиновые кислоты были обнаружены в 1868 г. швейцарским ученым Мишером (F. Miescher), который установил, что эти вещества локализуются в ядрах клеток, обладают кислотными свойствами и в отличие от белков содержат фосфор. Химически Н. к. являются полинуклеотидами, т.е. биополимерами, построенными из мономерных звеньев — мононуклеотидов, или нуклеотидов (фосфорных эфиров так называемых нуклеозидов — производных пуриновых и пиримидиновых азотистых оснований, D-рибозы или 2-дезокси-D-рибозы). Пуриновыми основаниями, входящими в молекулу ДНК, являются аденин (А) и гуанин (Г), пиримидиновыми — цитозин (Ц) и тимин (Т). В нуклеозидах РНК вместо тимина присутствует урацил (У). В полинуклеотидную цепь нуклеотиды соединяются посредством фосфодиэфирной связи (рис. 1).
Первичная структура Н. к. определяется порядком чередования азотистых оснований, а их пространственная конфигурация — нековалентными взаимодействиями между участками молекулы: водородными связями между азотистыми основаниями, гидрофобными взаимодействиями между плоскостями пар оснований, электростатическими взаимодействиями с участием отрицательно заряженных фосфатных групп и противоионов.
Дезоксирибонуклеиновые кислоты, выделенные из различных организмов, отличаются по соотношению входящих в их состав азотистых оснований, т.е. по нуклеотидному составу, который у всех ДНК подчиняется правилу Чаргаффа: 1) число молекул аденина в молекуле Н. к. равно числу молекул тимина, т.е. А = Т; 2) число молекул гуанина равно числу молекул цитозина, т.е. Г = Ц; 3) число молекул пуриновых оснований равно числу молекул пиримидиновых оснований; 4) число 6-аминогрупп равно числу 6-кетогрупп, что означает, что сумма аденин + цитозин равна сумме гуанин + тимин, т.е. А + Ц = Г + Т. Правило Чаргаффа справедливо и для так называемых минорных азотистых оснований (метилированных или других производных пуриновых и пиримидиновых оснований). Таким образом, нуклеотидный состав каждой ДНК характеризуется постоянной величиной — молярным соотношением  (фактором специфичности) или процентным содержанием Г—Ц-пар, т.е.
(фактором специфичности) или процентным содержанием Г—Ц-пар, т.е.  . Величина последнего показателя практически одинакова для организмов одного класса. У высших растений и позвоночных животных она составляет 0,55—0,93.
. Величина последнего показателя практически одинакова для организмов одного класса. У высших растений и позвоночных животных она составляет 0,55—0,93.
(фактором специфичности) или процентным содержанием Г—Ц-пар, т.е. . Величина последнего показателя практически одинакова для организмов одного класса. У высших растений и позвоночных животных она составляет 0,55—0,93. В 1953 г. американский биохимик Уотсон (J.D. Watson) и английский биофизик Крик (F.Н. Crick), основываясь на правиле Чаргаффа и данных рентгеноструктурного анализа молекул ДНК, установили, что молекула ДНК имеет вид двойной спирали, в которой две антипараллельно направленные углеводно-фосфатные цепи удерживаются водородными связями между аденином и тимином и гуанином и цитозином. Последовательность азотистых оснований в одной цепи определяет их последовательность в другой нуклеотидной цепи, т.к. размеры комплементарных пар А—Т и Г—Ц одинаковы, что позволяет нуклеотидной цепи свернуться в правильную двойную спираль, определяющую вторичную структуру ДНК. На один виток такой спирали приходится 10 пар оснований.
Генетическая информация «зашифрована» в молекуле ДНК в комбинациях нуклеотидов, кодирующих включение в строящуюся полипептидную цепь определенных аминокислот. Установлено, что генетический код универсален для всех живых существ; триплетен, т.е. каждая аминокислота кодируется тройкой нуклеотидов (триплетом); не перекрывается, т.е. данный нуклеотид может входить только в один так называемый кодон; одна аминокислота может кодироваться несколькими триплетами; большинство нуклеотидных триплетов имеет «смысл», т.е. кодирует аминокислоты.
Рентгеноструктурный анализ молекул ДНК показал, что пуриновые и пиримидиновые основания нуклеотида лежат в одной плоскости, практически перпендикулярной продольной оси молекулы (так называемая В-форма двойной спирали ДНК, которую она имеет в физиологических условиях), тогда как остатки дезоксирибозы находятся в плоскости, почти перпендикулярной плоскости, в которой лежат азотистые основания.
Синтез Н. к. в клетке осуществляется по принципу копирования молекулы-матрицы путем реакции поликонденсации нуклеозидтрифосфатов с отщеплением пирофосфата. Этот процесс катализируется ферментами полимеразами. Последовательность азотистых оснований в строящейся молекуле определяется их последовательностью в молекуле-матрице. Синтез молекул ДНК называют репликацией (редупликацией), т.е. образованием молекул-реплик на материнской молекуле.
Генетическая информация из клетки в клетку, из поколения в поколение передается именно путем репликации (редупликации) молекул ДНК, в результате которой из одной молекулы ДНК образуются две дочерние, полностью идентичные материнской, что обеспечивает передачу полного комплекса наследственной информации. В процессе редупликации между парами нуклеотидов разрываются водородные связи, к освободившимся нуклеотидам присоединяются содержащие комплементарные им азотистые основания дезоксинуклеозидтрифосфаты. Т.о., каждая дочерняя двойная спираль включает в себя одну материнскую и одну вновь синтезированную полинуклеотидную цепь. Репликация является сложным процессом, в котором принимают участие множество ферментов, белок, разделяющий нити ДНК, нуклеазы, лигазы и другие белки. Для ее осуществления необходимы матричная ДНК; дезоксирибонуклеозидтрифосфаты всех четырех азотистых оснований, а также ионы Mg2+ (рис. 2).
Рост новой цепи катализируется ферментом ДНК-полимеразой. Репликация начинается (инициируется) в определенных участках молекулы ДНК — репликаторах, первичная структура которых характеризуется высоким содержанием пар А—Т и наличием так называемых обратных повторов (палиндромов). Терминация (окончание) репликации происходит либо при слиянии двух вилок репликации, либо в так называемых точках терминации редупликации. У бактерий и эукариот в каждом цикле деления клетки, как правило, должна реплицироваться вся ДНК и при том один раз, поэтому должны существовать механизмы контроля за инициацией репликации и механизмы, благодаря которым различаются материнские и дочерние молекулы. Иногда (в норме и при патологии) может происходить многократная репликация ДНК всего генома (см. Ген) без последующего деления клетки, что приводит к возникновению полиплоидных ядер, или репликация отдельных частей генома без репликации всего генома (так называемая экстрарепликация). Описаны случаи недорепликации части ДНК той части генома в клетках эукариот, в которой нет генов, необходимых для жизнеобеспечения клетки. Сходство ферментов, катализирующих этапы репликации, и основных процессов, происходящих в вилке репликации, у прокариот и эукариот свидетельствует о высокой эволюционной стабильности и жестком генетическом контроле репликации ДНК.
Распад ДНК, как и РНК, в норме в живых клетках не происходит. ДНК погибших клеток или клеток, целостность стенки которых нарушена, расщепляется специфическими нуклеазами (ДНК-азой I и ДНК-азой II), катализирующими разрыв межнуклеотидных связей в поли- или олигонуклеотидах без образования неорганического фосфата. По характеру действия нуклеазы являются фосфодиэстеразами. Роль нуклеаз в генетической рекомбинации (см. Генетическая инженерия), исправлении (репарации) генетических повреждений, защите клетки от чужеродных Н. к. чрезвычайно велика. Их активность в тканях и биологических жидкостях может служить диагностическим тестом при ряде заболеваний. Так, активность ДНК-азы II в сыворотке крови возрастает при острых панкреатитах, особенно геморрагическом панкреатите, а активность РНК-азы в сыворотке крови возрастает при уремии. Генетически обусловленная недостаточность некоторых нуклеаз является причиной тяжелых наследственных заболеваний (например, пигментной ксеродермы).
По способу атаки субстрата нуклеазы делят на экзо- и эндонуклеазы. Экзонуклеазы катализируют последовательное отщепление моно- или олигонуклеотидов от одного из концов полинуклеотидной цепи. Некоторые экзонуклеазы катализируют расщепление и ДНК, и РНК.
Эндонуклеазы катализируют разрыв между внутренними звеньями полинуклеотидной цепи и отличаются гораздо более высокой субстратной специфичностью, чем экзонуклеазы.
В распаде Н. к. принимают участие также нуклеозидазы, которые катализируют расщепление фосфомоноэфирных связей в мононуклеотидах с образованием нуклеозидов и неорганического фосфата, по характеру действия они являются фосфомоноэстеразами, и гидролитические и фосфоролитические нуклеозидазы (нуклеозидгидролазы и нуклеозидфосфорилазы).
Рибонуклеиновые кислоты у большинства организмов обеспечивают реализацию генетической информации, однако у РНК-содержащих вирусов они могут быть также носителями наследственной информации подобно ДНК.
Молекула РНК представляет собой линейный полимер, мономерными звеньями которого являются рибонуклеотиды (их углеводным компонентом служит пентоза—D-1-рибоза). Характерные структурные элементы некоторых РНК представлены минорными основаниями (соответствующие им нуклеотиды входят в состав ряда РНК в очень небольших количествах).
Первичная структура РНК строго специфична и уникальна для каждого вида природной РНК. Она служит формой записи биологической информации, многократно и точно воспроизводящейся в процессе биосинтеза РНК. Структура синтезируемой РНК, строящейся на молекуле ДНК как на матрице (процесс транскрипции), определяется этой молекулой ДНК, что является начальным этапом реализации генетической информации, зашифрованной в ее полинуклеотидной цепи. Синтез РНК-транскрипция катализируется РНК-полимеразами, которые считывают лишь одну, так называемую значащую нить двойной спирали ДНК-матрицы. В процессе транскрипции образуется РНК-копия соответствующей ДНК или РНК-копия гена.
Вторичная и третичная структуры молекулы РНК (ее пространственная конфигурация), как и в молекулах ДНК, формируются в основном за счет водородных связей и межплоскостных гидрофобных взаимодействий между азотистыми основаниями. Однако, если для молекулы ДНК характерен вид устойчивой двунитевой спирали, то вторичная и третичная структуры молекул РНК гораздо более лабильны и вариабельны. Полинуклеотидные цепи РНК обладают большой гибкостью. В растворах с низкой ионной силой молекулы РНК ведут себя как типичные полиэлектролиты; при повышении ионной силы раствора разбухшие цепи РНК сжимаются, на отдельных участках гибкой цепи РНК, которая, перегибаясь, навивается сама на себя, образуются двуспиральные структуры в результате так называемого комплементарного спаривания, как и в молекулах ДНК. Такие структуры в молекулах РНК стабилизируются водородными связями между противолежащими азотистыми основаниями на антипараллельных участках цепи. Специфическими парами азотистых оснований на двуспиральных участках молекулы РНК являются, как и в молекуле ДНК, А—У, Г—Ц и Г—У (урацил вместо тимина). Молекулы РНК, состоящие из двух комплементарных полинуклеотидных цепей, обнаружены в некоторых вирусах; кроме того, они образуются как промежуточные формы при биосинтезе ряда вирусных РНК (так называемые репликативные формы РНК).
Некоторые двуспиральные РНК подобно ДНК могут существовать в виде кольцевых молекул и, если обе полинуклеотидные цепи ковалентно замкнуты, способны образовывать суперспирализованные кольца. РНК могут формировать двухтяжевые комплексы, в которых один тяж представлен полирибонуклеотидной, а другой — полидезоксирибонуклеотидной цепью. Такие ДНК—РНК-гибридные комплексы образуются во время репликации ДНК с участием так называемых затравочных фрагментов РНК, а также во время транскрипции РНК на матрице ДНК. ДНК—РНК-гибридные комплексы возникают также после заражения клеток некоторыми РНК-содержащими вирусами в результате синтеза на вирусной РНК комплементарной ей ДНК с помощью фермента обратной транскриптазы (ревертазы).
Содержание РНК в живых клетках (за исключением сперматозоидов) значительно выше, чем содержание ДНК. Основная масса РНК локализована в цитоплазме клетки: в составе собственно цитоплазматических рибосом (см. Клетка) и митохондрий, а также присутствует в виде нерибосомных комплексов с белками. В ядро РНК входит хроматин (часть ядерных РНК является продуктом текущей транскрипции генов).
Функции РНК в клетке сложны и многообразны. Различают три основных типа РНК: рибосомные РНК (рРНК), транспортные РНК (тРНК) и информационные, или матричные, РНК (иРНК, или мРНК). В клетках существует еще набор так называемых малых РНК, функции которых пока не выяснены.
Рибосомные РНК составляют около 80% всех клеточных РНК, по массе от 50 до 65% всего материала рибосом приходится на РНК. В каждой субъединице рибосом (большой и малой) РНК служит каркасом, на котором собираются рибосомные белки Сформировавшийся рибонуклеопротеидный комплекс (так называемый рибонуклеопротеидный тяж — РНП-тяж) организуется в сложную компактную частицу — собственно рибосомную субъединицу. Роль рРНК в белоксинтезирующей системе клетки не исчерпывается ее структурными функциями. Полагают, что некоторые участки рРНК играют определенную роль в формировании пептидилтрансферазного центра рибосомы, ответственного за образование пептидных связей между остатками аминокислот при синтезе белка.
Транспортные РНК составляют около 15% от общего количества клеточных РНК. Нуклеотидная цепь тРНК содержит всего 75—90 нуклеотидов, для большинства тРНК установлена полная последовательность нуклеотидов в цепи. Особенностью тРНК является относительно высокое содержание нуклеотидов, включающих минорные азотистые основания. Эти Н. к. с помощью высокоспецифичных ферментов аминоацил-тРНК-синтетаз присоединяют к себе ту или иную аминокислоту и переносят ее на рибосому. Для одной и той же аминокислоты имеется несколько тРНК, которые называют изоакцепторными. Транспортная РНК в ходе синтеза полипептидной цепи белка «узнает» специфическую аминоацил-тРНК-синтетазу, принимает от нее активированную аминокислоту, присоединяется к иРНК на рибосоме и тем самым обеспечивает строгую специфичность выбора и встраивания аминокислот в растущую молекулу белка; после образования пептидной связи между доставленной аминокислотой и уже построенной полипептидной цепью тРНК удерживает эту цепь на рибосоме.
Информационные, или матричные, РНК составляют всего 5% от общего количества клеточной РНК. Их биологическая роль заключается в программировании синтеза всех клеточных белков. Если рРНК и тРНК относятся к «обслуживающему» аппарату белоксинтезирующей системы клетки, то иРНК является прямым посредником между ДНК и белками и служит матрицей для синтеза белков; в форму иРНК переводится большая часть информации, заключенной в ДНК.
В соответствии с химическим строением полинуклеотидной цепи существуют три группы методов количественного определения Н. к.: по содержанию азотистых оснований (спектрофотометрическое определение), углеводного компонента (специфические цветные реакции), по количеству фосфора. Методы второй группы специфичны к типу Н. к. и позволяют отличить ДНК от РНК.
Определение нуклеотидных последовательностей индивидуальных Н. к. представляет большой практический интерес. Это, в частности, путь идентификации мутантных генов и расшифровки молекулярных механизмов, лежащих в основе синтеза аномальных белков при различных формах наследственной патологии (см. Наследственные болезни).
См. также Пиримидиновый обмен, Пуриновый обмен.
Библиогр.: Гайцхски В.С. Информационные РНК клеток животных. М., 1980; Ролан Ж.К., Селоши А. и Селоши Д. Атлас по биологии клетки, пер. с франц., с. 28, М., 1978; Спирин А.С. и Гаврилова Л.П. Рибосома, М., 1968; Уотсон Дж. Д. Молекулярная биология гена, пер. с англ., М., 1978; Шабарова З.А. и Богданов Л.А. Химия нуклеиновых кислот и их компонентов.
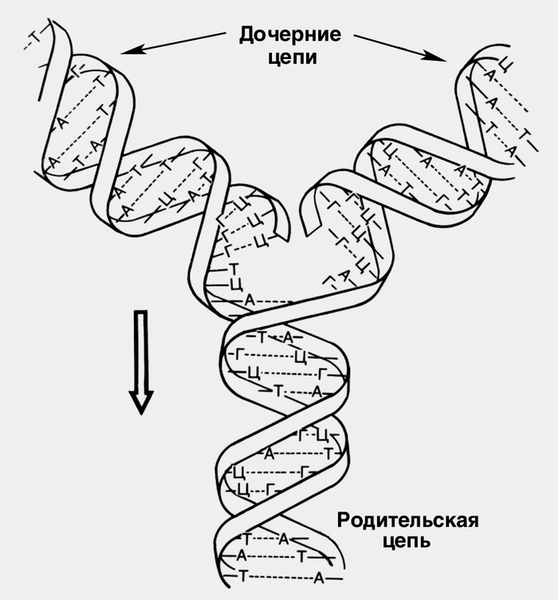 реплика) строится на каждой из родительских полинуклеотидных цепей, как на матрице. Стрелкой указано направление движения так называемой вилки репликации, пунктиром обозначены водородные связи между азотистыми основаниями. А — аденин, Т — тимин, Г — гуанин, Ц — цитозин">
реплика) строится на каждой из родительских полинуклеотидных цепей, как на матрице. Стрелкой указано направление движения так называемой вилки репликации, пунктиром обозначены водородные связи между азотистыми основаниями. А — аденин, Т — тимин, Г — гуанин, Ц — цитозин">Рис. 2. Схема репликации молекулы ДНК: дочерняя цепь (реплика) строится на каждой из родительских полинуклеотидных цепей, как на матрице. Стрелкой указано направление движения так называемой вилки репликации, пунктиром обозначены водородные связи между азотистыми основаниями. А — аденин, Т — тимин, Г — гуанин, Ц — цитозин.
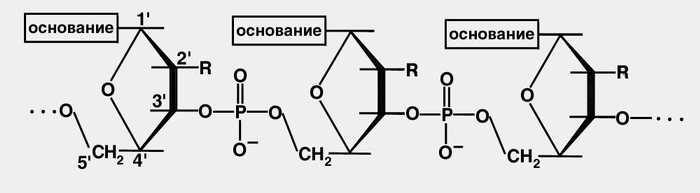
Рис. 1. Схематическое изображение соединения нуклеотидных звеньев в полинуклеотидную цепь нуклеиновой кислоты: в молекулах ДНК R-водород, в молекулах РНК — ОН-группа (гидроксильная группа).
1. Малая медицинская энциклопедия. — М.: Медицинская энциклопедия. 1991—96 гг. 2. Первая медицинская помощь. — М.: Большая Российская Энциклопедия. 1994 г. 3. Энциклопедический словарь медицинских терминов. — М.: Советская энциклопедия. — 1982—1984 гг.